一、前言
位於台灣東南方海面的蘭嶼,有許多獨特的動物相(fauna)及植物相(flora)。許多生物在型態(morphology)上與台灣的生物有顯著的差異,因而蘭嶼一直是野外觀察者所嚮往的島嶼。這個現象早在二十世紀初期日本學者大島正滿(Masamitsu Oshima)發現,並提出將華萊士線(Wallace’s Line)往北延伸的見解。而後有黑田長禮(Nagamichi Kuroda)等人以鳥類、哺乳類的觀點認為蘭嶼的生物相應屬於台灣的一部份。但是在1933年由鹿野忠雄(Tadao Kano)針對蘭嶼的甲蟲及蝸牛的觀點認為蘭嶼與台灣的動物相差異甚大,反而與綠島、巴丹島及呂宋島相似,提出華萊士線應北伸到台灣及蘭嶼及綠島之間,稱為新華萊士線(Neo-Wallace’s Line)。本篇就生物的分布、生物界線(biological boundary)的劃設及冰河時期海平面下降的現象與鹿野忠雄的研究做介紹與討論華萊士線的變遷。
二、華萊士線的變遷與背景
華萊士(Alfred R. Wallace)是英國的動物地理學者,為生物地理學之父。在十八世紀時於印度尼西亞(Indonesia)一帶做野外生物調查,分析陸生性動物在該地的動物相及群集(Community)分布。漸漸的在所蒐集的資料當中,反映出生物在全世界的分布狀況。其中最明顯的動物相差異(faunal gap)在峇里島(Bali)與龍目島(Lombok)之間。根據他的說法,印尼峇里島的動物系以亞洲的要素的動物呈優勢,而龍目島以東以新幾內亞(New Guneia)要素的動物呈優勢(山崎柄根2001)。」
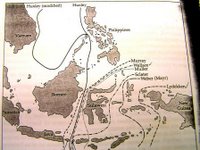
後來此線北伸到婆羅洲(Borneo)及蘇拉威西(Sulawesi)之間,並通過菲律賓(Philippine)群島西側。赫胥黎(T.H.Huxley)將華萊士線所引的動物相分界稱之為新華萊士線。
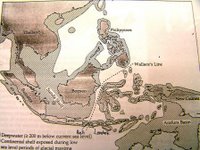
有趣的是,華萊士線與東南亞在更新世(Pleistocene)冰河時期陸地的(當時該陸塊稱為Sunda Shelf)。另有一條劃過新幾內亞及澳洲(Australia)海岸線一致西側的Lydekker’s line也與當時的陸地海岸線相合(該陸塊稱為Arafura Basin)。可見動物相的分布與海平面下降的的地理分布息息相關(圖一)。
E.D.Merrilly在二十世紀時在菲律賓群島及婆羅洲調查龍腦香科(Family Dipt-erocarpaceae)植物。他認為華萊士線應向北延伸到菲律賓群島西側(Palawan群島除外)並繞到呂宋島北側。此論點受到R.E.Dickerson支持。因而此修正線被稱為梅里爾線(Merrill’s Line)、新華萊士線或赫胥黎線(Huxley’s Line)(圖二)。
三、台灣蘭嶼的動物相差異及華萊士線的北伸
二十世紀初期日本學者大島正滿提出華萊士線北伸的可能性,再透過鹿野忠雄的實際調查發現蘭嶼的動物相與華萊士線有關,並從生物地理學的角度切入做進一步研究,再加上台灣高山冰蝕地形的發現來思考台灣與蘭嶼的生物地理關係。
起初黑田長禮、紉山米太郎、岡田彌一等人從蘭嶼島的鳥類、哺乳類、兩棲類及爬行動物的分布,認為蘭嶼的動物相應視為台灣的一部份。但是鹿野忠雄對於蘭嶼的甲蟲相注意到具有菲律賓動物相的特徵,因而質疑納入台灣動物相的說法。
鹿野忠雄在蘭嶼發現六種象鼻蟲,認為其中有蘭嶼特有種或分布於綠島,或屬於巴布煙(Babuyan)群島及呂宋島的亞種。由於這種象鼻蟲不具有飛行能力,步行移動緩慢,容易形成地理隔離。僅在呂宋島就有58種Pachyrrhynchus屬的象鼻蟲。另外蘭嶼光澤蝸牛(Helicostyla okadai)也僅分布於蘭嶼綠島地區,但不見於台灣,且同屬在呂宋島有95種。
因此,鹿野忠雄認為,此類物種容易形成地理隔離,如果透過海漂方式不論是成蟲或幼蟲,都很容易在抵達陸地之前死亡。所以在菲律賓群島、蘭嶼綠島之間的分布並非以海漂式散布,否則距離較遠的台灣應該有此類物種的分布。由此可知,牠們的分佈方式應該是以陸行方式散佈,而支持菲律賓群島與蘭嶼綠島曾經與陸橋相連的假說。由於氣候暖化、海平面上升以及蘭嶼綠島的火山作用重新將物種隔離起來,並各自在島上演化,形成現在的生物相。再加上鹿野忠雄在蘭嶼的動物相調查,因而認為華萊士線應北伸至台灣與蘭嶼綠島之間。
四、討論
在華萊士線的劃設過程中,最有趣的是華萊士線幾乎與冰河時期的陸地海岸線一致。華萊士的動物相資料反映出海洋是生物地理隔離的重要因素。
更新世冰河時期,海平面大約比現在低120公尺,當時中南半島與蘇門達臘(Sumatra)、印度尼西亞、婆羅洲皆相連;台灣與亞洲相連;新幾內亞與澳洲也有相連。而華萊士線劃過的地方如婆羅洲與蘇拉威西之間、台灣與蘭嶼之間、峇里島與龍目島之間仍有海洋的阻隔(圖三)。
以往認為,相距較近的陸地會有較相似的動物相。但是婆羅洲卻與中南半島較相似,反而與距離較近的蘇拉威西差異很大。因為中南半島與婆羅洲曾經相連,而大多數的動物以陸行方式散佈遷移,所以即使距離較近,大部分動物也無法跨過海洋的阻隔。
相較之下,某些較不受海洋阻隔影響的動物(如鳥類),我認為並不適用於整體動物地理分布的研究。因為其行動力較大較自由,而且一旦遷徙常常就是繞過大半個地球,分布過於廣泛。應該針對行動能力低、容易受隔離的物種作研究,例如無脊椎動物、軟體動物、植物等等。當然綜觀該地區所有的物種分布再做分析判斷更是必要的。所以我認為當時黑田長禮等人的說法並不具足夠的說服力。而且在台灣與蘭嶼的鳥類相,有些物種的確有特殊的差異。例如綬帶鳥(Terpsiphone atrocaudaat atrocaudata)與黑綬帶鳥(Terpsiphone atrocaudaat periophthalmica )、藍磯鶇(Monticola solitarius)與藍腹藍磯鶇(Monticola solitarius pandoo)、綠繡眼(Zost- erops japonica)與低地繡眼(Zosterops meyeni)等。但這又牽扯到島間的距離,鳥類是否能離開棲地太久等因素。
因此,冰河時期的陸地相連是動物分布遷移的一大時機,同時海洋也限制動物的分布範圍,且這些會顯現在地區與地區之間的生物相差異上。
五、結語
事實上,生物相的調查並不容易,再加上生物相的複雜多變,影響生物相變化的因素又很多:海平面也許又會變動、陸地板塊不斷的在推擠漂移、生命不斷的在演化、種化、絕種。生物相只會更加的複雜、更難以研究。所以要在這模糊複雜的生物相畫上一條清晰的界線,本來就是一件矛盾的事情。而且生物相能不斷的變動,這些生物界線還會跟著不斷的修改、變遷。
所以,生物界線只能給我們一個整體性的概念,並不能給予最確切的答案。雖然大自然有規則,但是沒有鐵律,所有的規則都不是一成不變的,都有例外存在。也提醒自己能銘記著,自然科學不是一個鑽牛角尖的學門,因為大自然千變萬化。同時,這也是自然科學如此吸引人的原因。
六、參考文獻
Harold K. Voris. 2000. Maps of Pleistocene sea levels in Southeast Asia : shore lines, river systems and time durations. Chicago. USA.
Times Book. 1985. The Time ATLAS of the World. 7th comprehensive edition.
James H. Brown & Mark V. Lomolino. 1998. Biogeography. second edition. USA.
山崎柄根著,楊南郡譯註。2001。鹿野忠雄-縱橫台灣山林的博物學者。晨星出版有限公司。台中。台灣。
林文宏。1997。台灣鳥類發現史。玉山出版股份有限公司。台北。台灣。
鹿野忠雄、鷹司信輔。昭和9年。蘭嶼的鳥相。日本鳥學會。日本。
顏重威。1998。台灣的野生鳥類(留鳥篇)。渡假出版社。台北。台灣。
吳永華。1996。被遺忘的日籍動物學者。晨星出版有限公司。台中。台灣。
七、圖片來源
圖一:Harold K. Voris. 2000. Maps of Pleistocene sea levels in Southeast Asia :shore lines, river systems and time durations.Chicago. USA.
圖二:James H. Brown & Mark V. Lomolino. 1998. Biogeography. second edition. USA. pp310.
圖三:James H. Brown & Mark V. Lomolino. 1998. Biogeography. second edition. USA. pp187.



